












 |
Владимир Емельянович Харин (1957-2013)
Кандидат биологических наук, старший научный сотрудник Лаборатоии ихтиологии Института биологии моря им. А.В. Жирмунского ДВО РАН, являлся единственным специалистом по морским рептилиям в России. Используя богатейшие коллекции Тихоокеанского института рыбного хозяйства и океанографии (ныне большая их часть хранится в Зоологическом музее Дальневосточного государственного университета, г. Владивосток), за 25 лет работы он описал 7 новых для науки видов морских змей (всего в мировой фауне известно 63 вида). В.Е. Харин провел ревизию Hydrophiidae s.l., в результате чего были установлены 2 новых подсемейства, 1 триба, 1 новый род (Pseudolaticauda) и 2 новых подрода, которые некоторыми специалистами рассматриваются в ранге родов. Наконец, им было обосновано разделение морских змей на 2 семейства – Laticaudidae (морские крайты) и Hydrophiidae (собственно морские змеи).
Редактор Алексей Викторович Чернышев, доктор биологических наук, ведущий научный сотрудник Лаборатории эмбриологии Института биологии моря им. А.В. Жирмунского ДВО РАН.
Перевод текста на английский язык осуществлен Натальей Владиславовной Мирошниковой.
Техническое редактирование текста выполнено Верой Сенкиновной Мун.
Разработчик сайта Виталий Олегeович Пойс.
Ввод текста и изображений - Лариса Георгиевна Ерофеева.
- Автор: admin
- Просмотров: 720
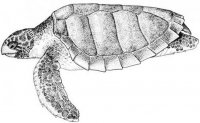
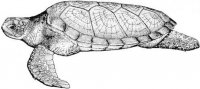
В российских водах Японского моря морские рептилии отмечены с конца XIX в. Прежде всего это находки морских змей – двухцветной пеламиды Pelamis platura в зал. Петра Великого (зал. Посьета в 1873 и 2006 гг.) (Strauch, 1874; Kharin, 2007; Харин, 2008) и большого морского крайта Pseudolaticauda semifasciata (у мыса Соснового в 1978 г.) (Чугунов, 1980; Харин, 2008). Кожистая черепаха Dermochelys coriacea была поймана в зал. Рында (1936 г.) и зал. Петра Великого: между островами Аскольда и Путятина (1972 г.), в бухтах Астафьева (1979 г.) и Спасения (1979 г.), у мыса Поворотного (1984 г.) и у западного побережья о-ва Сахалин в районе пос. Правда Невельского района (Емельянов, 1937; Братчик, Воронцов, 1974; Благодеров, 1982; Боркин, Басарукин, 1986). Единственный экземпляр логгерхеда Caretta caretta был пойман в бух. Маньчжур зал. Петра Великого (1940 г.) (Терентьев, Чернов, 1949; Благодеров, 1982).
Автор первой находки кожистой черепахи предполагал, что эта поимка, вероятно, связана с прохождением у берегов Приморья ветви теплого течения Соя (Емельянов, 1937). Если же сопоставить годы поимок морских змей и морских черепах в российских водах Японского моря с динамикой численности дальневосточной сардины-иваси Sardinops melanosticta, то выявляются, по-видимому, неслучайные параллели. Благодаря исследованиям Шунтова и Василькова (1981, 1982) установлено, что динамика уловов этого вида в дальневосточных морях связана с циклическими изменениями климата и температуры морской воды. Подъемы численности иваси в прошлом столетии приходились на периоды заметного потепления. В дальневосточных морях было отмечено продвижение к северу большого числа тропических и субтропических видов рыб (Шунтов, 1966). К этим же периодам в основном приурочены и находки морских рептилий (Боркин, Басарукин, 1986). С этой позиции сложно понять единственную находку в водах Приморья Caretta caretta в 1940 г., в период некоторого похолодания. Все же периодическое и участившееся с 70-х годов прошлого века появление морских рептилий в морях юга Дальнего Востока России, по-видимому, обусловлено периодическим потеплением климата и морской воды. Данное предположение также подтверждается систематическими находками тропических и субтропических видов рыб в это же время (Румянцев, 1947, 1951; Новиков, 1957; Маркевич, 2001, 2002б, 2005; Соколовский, Соколовская, 2004; Харин, Чеблуков, 2005; Харин, Милованкин, 2005; Харин, Маркевич, 2006; Харин и др., 2006, 2007; Долганов и др., 2007, 2008).
Существует также предположение, что заход в наши воды морских змей и черепах из южной части Японского моря, возможно, связан с глобальным потеплением (Харин, 2008). В таком случае новые находки морских рептилий представляются вполне вероятным событием.
- Автор: admin
- Просмотров: 738
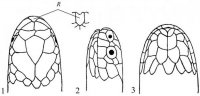
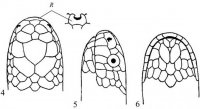
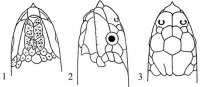
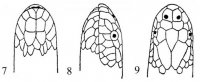
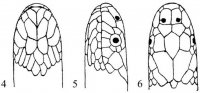
Морфология. Тело округлое или овальное, заключено в более или менее развитый костный панцирь. Панцирь (рис. I, 5, 6; рис. II, 1, 2) снаружи покрыт кожей или роговыми щитками совместного текального и эпитекального происхождения (Völker, 1913; Romer, 1956; Черепанов, 1988, 1996, 2002), связанными с ребрами и остистыми отростками спинных позвонков. Ребра причленены интервертебрально и охватывают снаружи пояса конечностей. Карапакс обычно состоит из следующих костей. По средней линии идет ряд непарных пластинок, первая из которых называется загривковой (nuchale); за нею следуют восемь невральных (neuralia), сросшихся с остистыми отростками спинных позвонков, и одна–три надхвостовых (suprapygalia) пластинок; последняя носит название хвостовой (pygale).
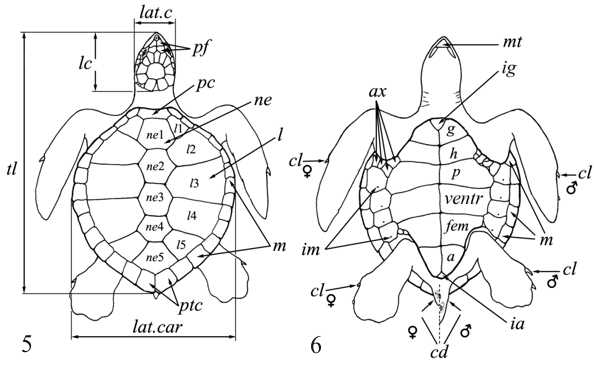
Рис. I, 5, 6. Строение морских черепах: 5 - вид сверху; 6 - вид снизу. Щитки: a – анальные, ax – подмышечные, f – лобный, fem – бедреные, fr– скуловой, g – горловые, h – плечевые, ia – межанальный, if – челюстные, ig– межгорловой, il – нижнегубные,im – подкраевые, l – реберные, m – краевые, mt – подбородочный, n – носовой, ne – позвоночные,p – грудные, par – теменной, pc – загривковый, pf – предлобные, po – предглазничные, pst – заглазничные, ptc – надхвостовые, r – межчелюстной, sl– верхнегубные, so– надглазничный, t – височные, ventr – брюшные. Другие обозначения: cd – хвост,cl – когти, lat.c – ширина головы, lat.car – ширина карапакса, lc – длина головы,tl – длина тела
По обе стороны от медиального ряда расположена серия парных реберных пластинок (costalia), сросшихся с ребрами, обычно также в числе восьми. Наружный край карапакса образован 11 парами краевых пластинок (marginalia). У большинства черепах карапакс покрыт роговыми щитками, границы между которыми отмечены бороздками на костных пластинках. Среди них различают медиальный ряд из загривкового (nuchale) и пяти позвоночных (vertebralia) щитков. С обеих сторон от медиального ряда лежат 4–5 пар реберных (costalia) щитков. По краю расположены 12 пар краевых щитков (marginalia). Иногда последняя пара срастается в один непарный хвостовой щиток (pygale). Пластрон состоит из девяти костных пластинок – непарного энтопластрона (entoplastron) и лежащих впереди него парных эпипластронов (epiplastron) и сзади тоже парных гиопластронов (hyoplastron), гипопластронов (hypoplastron) и ксифипластронов (xiphiplastron). Энтопластрон в некоторых случаях может отсутствовать. Передняя и задняя выемки в панцире, ограниченные краями карапакса и пластрона, получили названия подмышечной (incisuraaxillaris) и паховой вырезок (incisurainguinalis). Пластрон покрыт следующими парными роговыми щитками: горловыми (gularia), плечевыми (humeralia), подкраевыми (inframarginalia), грудными (pectoralia), брюшными (abdominalia), бедренными (femoralia) и анальными (analia). В ряде случаев впереди и между горловыми щитками помещается парный или непарный межгорловой щиток (intergulare). Панцирь полуводных и наземных черепах обычно остается полным, но у большинства водных и особенно морских видов он более или менее сильно редуцирован, причем тем сильнее, чем дальше зашла специализация: теряется костная связь между карапаксом и пластроном, которые соединены теперь лишь связками; ослабевают связи между отдельными костными элементами, между ними появляются фонтанели.
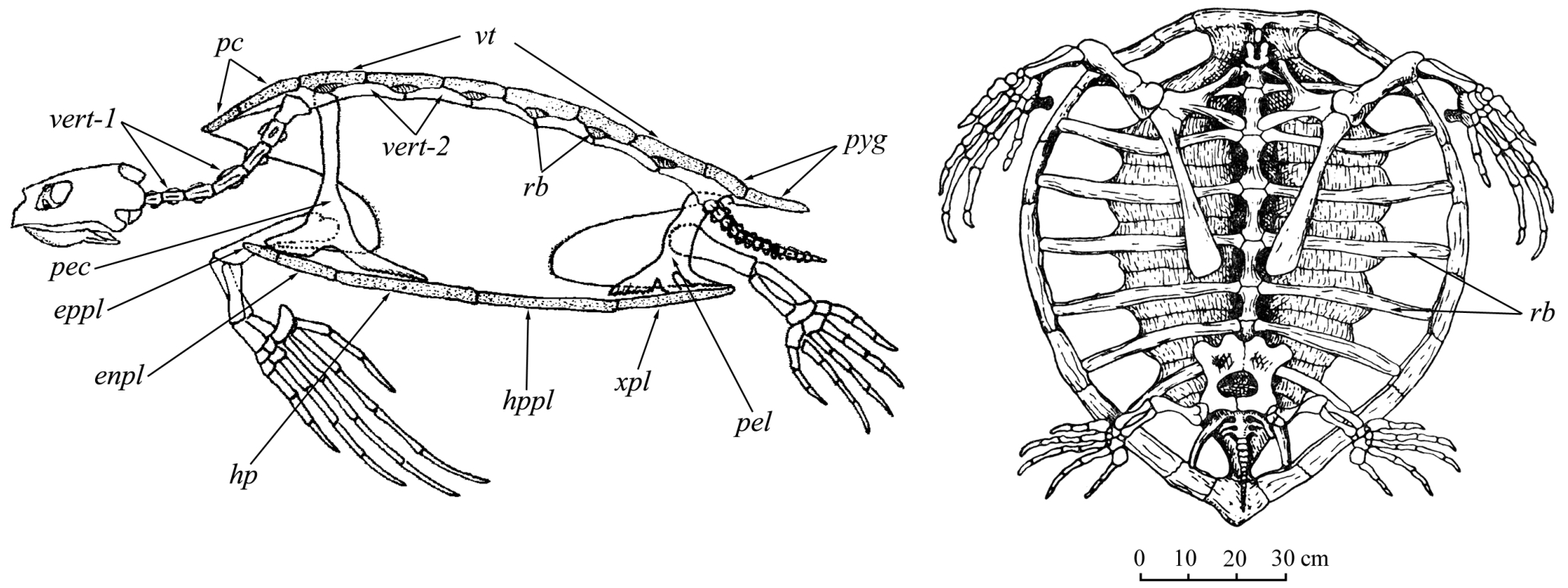
Рис. II, 1, 2. Общий вид скелета Chelonia mydas: 1- вид сбоку; 2 - вид снизу: enpl – эндопластрон, eppl – эпипластрон, hp – гиопластрон, hppl – гипопластрон, np – загривковая пластинка, pec – грудной пояс, pel – тазовый пояс, pyg – хвостовая пластинка, rb – ребра, vert-1 – шейные позвонки, vert-2 – позвонки тела, vt – позвоночные пластинки, xpl – ксифипластрон.
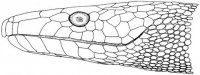
Череп современных черепах монолитно построен и полностью акинетичен (рис. III, 1, 2), элементы верхней челюсти, неба и дерматокраниума (dermatocranium) прочно связаны друг с другом и с нейрокраниумом (neurocranium).
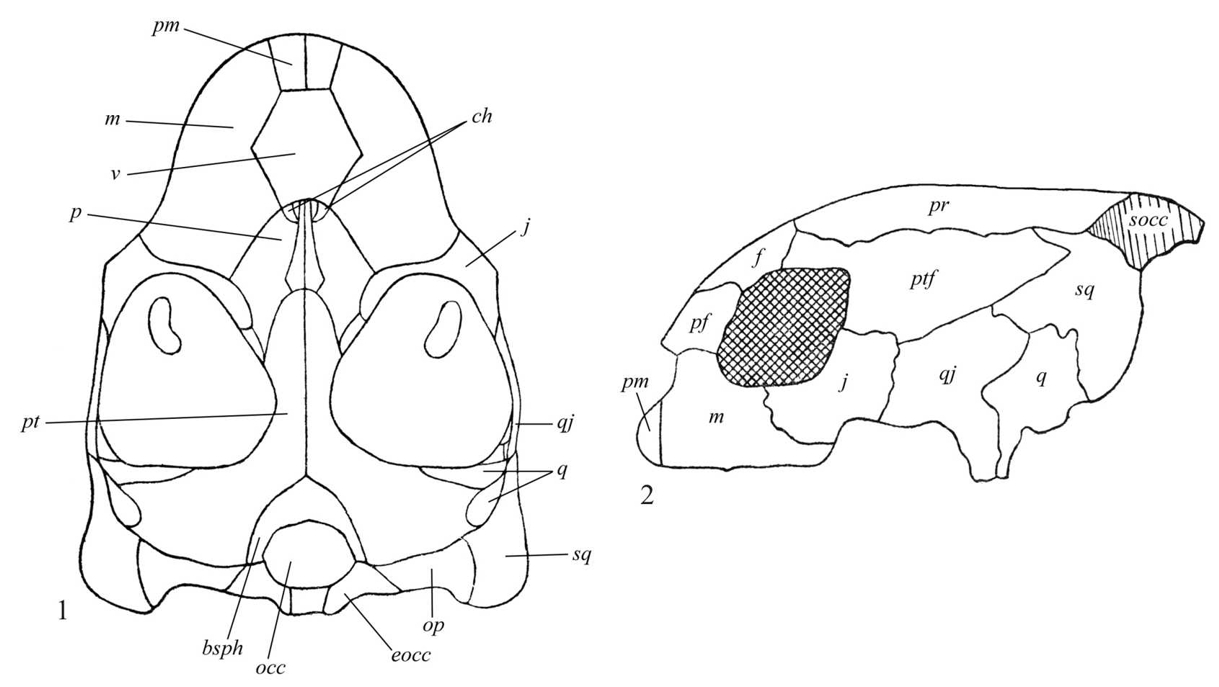
Рис. III, 1, 2. Череп морской черепахи. 1 - вид снизу; 2 - вид сбоку: bsph – базисфеноид, ch – отверстия хоан, eocc – боковая затылочная кость, f – лобная кость, j – скуловые кости, m – челюстная кость, occ – затылочная кость, op– заднеушная кость, p – небная кость, pf – предлобная кость, pm – межчелюстная кость, pr – теменная кость, pt – крыловидная кость, ptf – заднелобная кость, q – квадратная кость, qj – квадратноскуловая кость, socc – верхняя затылочная кость, sq – чешуйчатая кость, v – сошник
Он без височных окон (анапсидный тип черепа), с выпуклой крышей и верхнезатылочным гребнем (cristasupraoccipitalis). Череп характеризуется следующими чертами: лицевой отдел сильно укорочен; глазницы (orbitae) большие; зубы отсутствуют и функционально замещены роговым чехлом (клювом, rostrum). Височная область дерматокраниума гимнокротафического типа, т.е. в той или иной степени редуцирована посредством развития задней и латеральной вырезок, между которыми обычно сохраняются костные пряжки (образованные скуловыми, квадратно-скуловыми и чешуйчатыми костями). У некоторых видов редуцированы и эти пряжки, а задние височные вырезки соединены с латеральными (Romer, 1956). Покровные элементы крыши черепа склонны к редукции. Для челюстного аппарата этих животных характерна высокая жесткость, прочное крепление первичной верхней челюсти к черепной коробке, в чем выражается адаптация к высоким нагрузкам при обработке корма. Заднетеменные, надвисочные, заднелобные и таблитчатые кости отсутствуют. Теменного отверстия (foramenparietale) нет. Наружные ноздри (naresexterni) почти всегда слиты и расположены на конце рыла. Внутренние ноздри, или хоаны (choanae), находятся в передней части небного комплекса. Позади небный комплекс с обеих сторон отделен от края крыши подвисочными впадинами (fossaesubtemporales). Носовые кости присутствуют только у наземных форм, у морских черепах отсутствуют. Предлобные кости соединяются по средней линии. Слезные кости отсутствуют. Глазницы ограничены предлобными, челюстными, скуловыми, заглазничными и лобными костями. Череп полностью акинетичен (костные элементы небно-квадратного хряща (palatoquadratums. pterygoquadratum) сращены с мозговой коробкой). Межптеригоидные (interpterygoidea) ямы отсутствуют. Сошник непарный. Наружные крыловидные кости, за редким исключением, отсутствуют. Квадратные кости сращены медиально с сильно расширенными латеральными стенками ушных капсул и характерно изогнуты вокруг слуховых косточек, образуя воронковидную барабанную полость. Челюстной сустав расположен впереди затылочного мыщелка. Заднеушные кости не сращены с боковыми затылочными. Внутреннее ухо связано системой наполненных перелимфой полостей с пространством между боковой стенкой ушной капсулы и квадратной костью. В результате сильного увеличения слуховой капсулы происходит значительное преобразование пространства, занятого челюстной мускулатурой. Эти мышцы уже не ориентированы в исходном вертикальном направлении, а изгибаются над слуховой капсулой и отклоняются назад по направлению к задневисочным ямам, причем у всех продвинутых черепах они проходят поверх блоковидной структуры, называемой трохлеарным отростком. Однако в двух современных подотрядах эта структура развивается по-разному. У Cryptodira она образуется на передней поверхности слуховой капсулы, а у Pleurodira – в виде латерального отростка крыловидной кости (Gaffney, 1979). У Cryptodira крыловидная кость соединена швом с основной клиновидной и направлена наружу в сторону квадратной; надкрыловидная кость (suprapterygoideum) всегда имеется. У Pleurodira до основания мозговой коробки доходит медиальный вырост квадратной кости; надкрыловидная кость всегда отсутствует. Верхние крыловидные кости включены в стенку мозговой коробки. Зубные кости большие, занимают преобладающую часть наружной поверхности челюсти. Угловые кости маленькие. Парасфеноид у всех черепах (кроме Dermochelys, в черепе которого остается много хряща) сращен с базисфеноидом в единое целое. Базисфеноид с затылочным мыщелком. По бокам последнего располагаются боковые затылочные кости (occipitalialateralia). У большинства черепах (кроме отдельных Testudinidae) небо почти плоское. У многих семейств отмечена тенденция к образованию вторичного неба. У морских черепах в формировании вторичного неба принимают участие небные кости.
Наличие у черепах панциря определяет неподвижность части позвоночного столба (рис. II, 1-4).
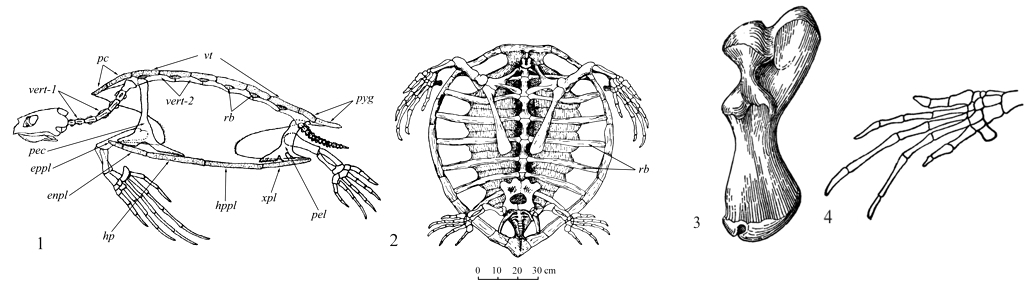
Рис. II, 1-4. 1 - общий вид скелета Chelonia mydas (вид сбоку): enpl – эндопластрон, eppl – эпипластрон, hp – гиопластрон, hppl – гипопластрон, np – загривковая пластинка, pec – грудной пояс, pel – тазовый пояс, pyg – хвостовая пластинка, rb – ребра, vert-1 – шейные позвонки, vert-2 – позвонки тела, vt – позвоночные пластинки, xpl – ксифипластрон; 2 - вид снизу; 3 - правая плечевая кость Chelonia mydas (по: Суханов, 1964); 4 - передняя левая конечность Caretta caretta (по: Romer, 1956).
Подвижны лишь хвостовой отдел и особенно шейный. Предкрестцовых позвонков 18 (8 шейных и 10 спинных), в крестце обычно 2 позвонка. Хвост относительно короткий. Шейные ребра отсутствуют. Спинные ребра, так же как и невральные дуги второго–девятого спинных позвонков, сращены с карапаксом. Грудины нет. Ключицы (claviculae) и межключицы (interclaviculae) включены в состав пластрона. Клейтрум (cleitrum) отсутствует. Первичный плечевой пояс трехлучевого строения, без прокаракоида (procaracoideum). Тироидные окна таза слиты с запирательными отверстиями и обычно разделены вентрально хрящом или костью. Плечевая (humerus) (рис. II, 3) и бедренные (femur) кости с полусферическими проксимальными головками, расположенными под прямым углом к длинной оси кости. Лучевая кость (radius) широкая и плоская, локтевая (ulna) – широкая, со слабо развитым локтевым отростком. У морских черепах и плечо, и предплечье располагаются в горизонтальной плоскости, но кости предплечья скрещиваются, как и у типичных наземных тетрапод. Однако в связи с тем что локоть у морских черепах обращен вперед, у них в проксимальной части предплечья локтевая кость находится снаружи, лучевая – внутри, а в дистальной – наоборот, тогда как ладонная поверхность обращена не вниз, а вверх. Движения в запястье и в пясти очень ограничены, и вся конечность превращена в мощный веслообразный плавник. При передвижении по суше морские черепахи опираются на плечо и на проксимальный край плавника.
Мыщелки челюстных суставов на квадратных костях у видов подотряда Cryptodira имеют относительно плоскую сочленовую поверхность, обращенную вентрально и несколько вперед; латеральный и медиальный бугры мыщелка разделены сужением кости и желобком, но слабовыпуклые. Соответствующие сочленовые ямки на нижней челюсти, образованные сочленовыми костями, у криптодир также почти плоские (со слабым срединным валиком), открытые (без резко выраженных краевых гребней), обращены дорсально и назад. Их длина в 2,0–2,5 раза превышает длину суставных поверхностей челюстных мыщелков квадратных костей. Такое устройство челюстных суставов позволяет нижней челюсти, помимо обычных дорсально-вентральных поворотов, совершать также продольные движения в пределах, допускаемых различиями длины суставных поверхностей верхней и нижней челюстей. У видов подотряда Pleurodira сочленовые поверхности мыщелков квадратных костей вогнутые, а суставных ямок нижней челюсти – выпуклые (Gaffney, 1975, 1979; Иорданский, 1990).
В связи с невозможностью дыхания реберного типа возник ряд приспособлений. У большинства видов колебания подъязычного аппарата создают вакуум в ротовой полости. Дыханию помогает движение конечностей и шеи. При вдохе происходит увеличение внутреннего объема тела, осуществляемое за счет изменения кривизны участков мягкой кожи, которая затягивает вырезки панциря и подстилается тонкими мышцами. При выдохе легкие сжимает окружающая их пленка, подтягиваемая специальными (диафрагмальными) мышцами. Находясь в защитной позе (с втянутыми в панцирь конечностями), черепаха дышать не может, поэтому длительное использование такой защиты требует устойчивости к гипоксии, так же как и пребывание под водой водных черепах.
Легкие имеют большой объем и сращены с внутренней выстилкой карапакса. Дополнительное водное дыхание у морских черепах осуществляется через выросты глотки, пронизанные кровеносными сосудами (Boulenger, 1889; Терентьев, 1961). У многих водных форм имеются анальные пузыри (bursaeanalis) – боковые тонкостенные выросты задней кишки непосредственно около клоаки, стенки которых обильно снабжены сосудами. Эти пузыри периодически наполняются водой.
В ротовой полости помещается толстый мясистый язык, широкая глотка ведет в пищевод, который постепенно переходит в длинный толстостенный желудок. Последний явственно отделен от кишечника кольцевым валиком. Двухлопастная печень и желчный пузырь имеют большие размеры, слепая кишка отсутствует. Мочевой пузырь большой.
Головной мозг черепах развит слабо и составляет меньше тысячной доли веса тела. Спинной мозг имеет значительную толщину и вес. Глаза хорошо развиты, зрачок всегда круглый, имеются два подвижных века и мигательная перепонка. У морских черепах в глазнице находится солевая железа, контролирующая отдачу жидкости и баланс солей. Слух развит слабо, барабанная перепонка у наземных черепах толстая, у морских слуховой проход замкнут сильно утолщенной кожей. Вкус, обоняние и осязание у черепах очень тонкие. Даже прикосновение к панцирю тотчас улавливается животным. Половая система самок представлена парными гроздевидных яичниками. У самцов черепахи имеется непарный копулятивный орган (penis), скрытый в клоаке и выдвигаемый при спаривании.
- Автор: Харин Владимир Емельянович
- Просмотров: 880
Пресмыкающиеся, птицы и млекопитающие составляют единую группу Amniota, отличающуюся от земноводных способом размножения, не зависящим от водной среды. Предполагается, что амниоты, по-видимому, составляют монофилетическую группу, произошедшую от одной из ветвей примитивных четвероногих в раннем карбоне. Известно, что группы, традиционно объединяемые в класс пресмыкающиеся, с филогенетической точки зрения неоднородны. У крокодилов гораздо более поздние общие предки с птицами, чем с черепахами или лепидозаврами (Антипчук, 1977). Предков млекопитающих обычно относят к пресмыкающимся, однако филогенетически они ближе к самим млекопитающим, чем к любому из современных подклассов рептилий. Исходя из этого, выделяют две группы амниот: млекопитающие вместе с их предками (Synapsida), с одной стороны, и все остальные – с другой. Последних, в свою очередь, также можно отнести к двум группам: это черепахи вместе со своими непосредственными предками (Anapsida), и линия, давшая начало лепидозаврам, крокодилам и птицам (Кэрролл, 1992, 1993). Термином “пресмыкающиеся” можно неформально обозначать черепах, крокодилов и лепидозавров,но указать точные границы класса Reptilia, за исключением, что это все амниоты, кроме птиц и млекопитающих, нельзя.
В последнее время Reptilia иногда рассматривают как монофилетическую группу, включающую всех амниот, за исключением Synapsida и их предков. При этом в составе рептилий выделяются две группы – Anapsida и Romeriida (включает диапсидных рептилий, а также птиц) (Gauthieretal., 1988).
- Автор: Харин Владимир Емельянович
- Просмотров: 735
Класс Reptilia в традиционном понимании – парафилетическая группа, в составе которой обычно выделяют следующие современные подклассы и отряды: подкласс Chelonia (черепахообразные) с единственным рецентным отрядом Testudines (черепахи); подкласс Lepidosauria (лепидозавры) с рецентными отрядами Sauria (= Lacertilia) – ящерицы, Serpentes (= Ophidia) – змеи и Rhynchocephalia – клювоголовые; подкласс Archosauria – архозавры с единственным рецентным отрядом Crocodilia (крокодилы) (Zugetal., 2001; Meyer, Zardoya, 2003). Ряд герпетологов рассматривает амфисбен (Amphisbaenia) как отдельный отряд лепидозавров (Bedriaga, 1884; Zangerl, 1944; Gans, 1978).
Некоторые авторы объединяют черепах вместе с представителями вымершего отряда Cotylosauria (котилозавры) в ранге подкласса Anapsida (Даревский, Орлов, 1988; Ромер, Парсонс, 1992; и др.). Тем не менее отнесение черепах к анапсидам спорно, поскольку их предки могли обособиться от примитивных капториноморф (Captorhinomorpha) позже синапсид или диапсид. К тому же их общее строение и образ жизни столь отличаются от присущих ранним анапсидам, что логичнее считать их отдельным подклассом (Siebenrock, 1897; Суханов, 1964; Gaffney, 1979; Кэрролл, 1992). Ивахненко (1984) объединяет большинство групп котилозавров, исключая капториноморф в узком смысле слова, с сеймуриаморфами (Seymouriamorpha) и черепахами в особый класс Parareptilia, параллельный настоящим рептилиям.
- Автор: Харин Владимир Емельянович
- Просмотров: 623