

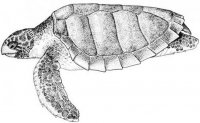

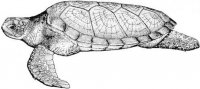



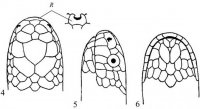



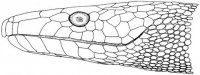




Морфология. Безногие лепидозавры с платибазальным черепом и подвижной квадратной костью, утратившие обе височные дуги. Межглазничная перегородка редуцирована или отсутствует. Глаза нормальные или редуцированы (подотряд Scolecophidia). Передняя часть неврокраниума (neurocranium) окостеневает и образована нисходящими отростками лобных костей. Подвижность черепа развита на границе носовых костей. Нисходящие отростки теменных костей срастаются с передней ушной костью (рис. I, 4).
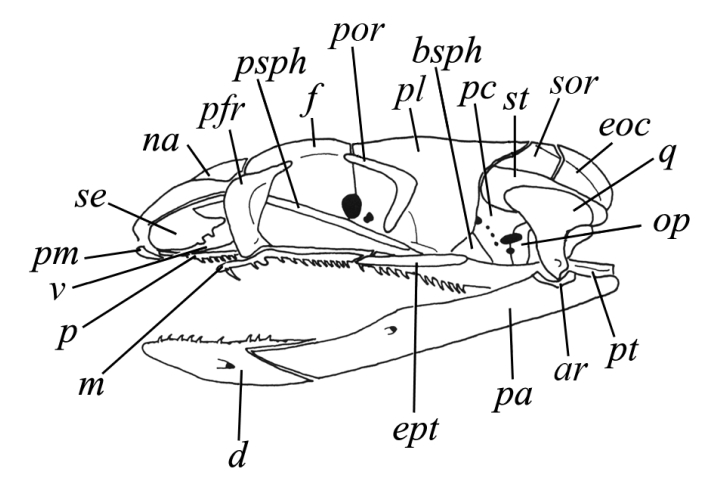
Рис. I, 4. Череп змеи. Кости: ar – сочленовая, bsph – базисфеноид, d – зубная, eoc – боковая затылочная, ept – наружная крыловидная, f – лобная, m – верхнечелюстная, na – носовая, op – задняя ушная, p – небная, pa – предсочленовая, pc – переднеушная, pfr – предлобная, pl– затылочная, pm – предчелюстная, por – заглазничная, psph – парасфеноид, pt – крыловидная, q – квадратная, se – покровная, sor – надглазничная, st – надвисочная, v – сошник.
Предчелюстные и, как правило, теменные кости непарные, теменного отверстия нет. Слезная, скуловая, чешуйчатая и обычно заднелобная кости утрачены. Лобная кость почти всегда достигает глазницы. Боковые затылочные кости смыкаются над затылочным отверстием. Надвисочная кость расположена на черепе поверхностно, ее задний конец часто выступает назад за уровень затылка и оказывается единственным элементом, к которому причленяется верхний конец квадратной кости. Квадратная кость значительно удлинена. Она подвижная (стрептостилия). Это преобразование способствует более широкому раскрыванию рта и связана с питанием крупной добычей (Иорданский, 1990; Zug etal., 2001). Базиптеригоидные отростки и верхняя крыловидная кость утрачены. Крыловидные кости теряют связь с мозговой коробкой, иногда они оканчиваются позади свободно, не соединяясь с квадратной костью. Небные и крыловидные кости подвижно соединяются с челюстной. Челюстная кость часто теряет связь с предчелюстной и становится подвижной, предчелюстная кость может редуцироваться. Ветви нижней челюсти соединены почти всегда связкой, задние кости нижней челюсти срастаются, самостоятельными остаются лишь зубная, пластинчатая и иногда венечная кости. Передняя половина нижней челюсти может становиться подвижной по отношению к задней. Зубы акродонтные, длинные, тонкие, изогнутые назад. Кроме челюстей зубы также присутствуют на крыловидных и небных костях. Замена зубов происходит постоянно по мере снашивания или утраты (полифиодонтия). Часто в передней (серии Proteroglypha, Solenoglypha) или задней части верхней (серия Opistoglypha) челюсти развиты длинные бороздчатые ядовитые клыки. Передние ядовитые клыки могут быть трубчатыми, пронизанными выводным каналом ядовитой железы. Во рту змей кроме межчелюстной (glandula intermaxillares), небных (glandula epalatinae), подъязычных (g. sublinguales) имеются еще верхние (g. labiale superiores) и нижние губные (g. l. inferiores) железы. Именно две последние становятся ядовитыми. Они находятся в связи с ядовитыми клыками, через желобок или канал которых происходит излияние ядовитого содержимого в рану благодаря сокращению жевательной мышцы (m. masseter).
Для большинства семейств змей характерен беспрецедентно широкий размах продольной подвижности, независимой для правой и левой половинок (гиперкинетизм). Эта подвижность используется при заглатывании добычи. Половинки челюстного аппарата работают поочередно: одной змея втаскивает добычу в глубь зева, а другая в это время раскрывается и выносится вперед для нового захвата (Pough, 1983). У ядовитых змей с несколько меньшей по размаху протракцией обеспечивается поворот качающейся верхнечелюстной кости для приведения сидящего на ней ядовитого зуба в рабочее положение. У примитивных семейств встречается мезокинетизм, их черепная коробка имеет гибкую зону в области глазниц. Слуховая косточка тонкая и прямая, идет от овального окна слуховой капсулы к квадратной кости; барабанной перепонки, полости среднего уха и евстахиевых труб нет. Внутренняя часть слуховой косточки погружена в вырост перилимфатического протока, заключенного в костный карман, образованный стенками слуховой капсулы. Подъязычная кость с одной жаберной дугой.
Позвонки процельные, укороченные, обычно сужающиеся кзади, с зигосфенами и зигантрами. По крайней мере передние туловищные позвонки несут также вентральные отростки – гипапофизы; часто развиты также крыловидные отростки, сидящие по сторонам невральной дуги. Гемальные дуги прирастают к телам позвонков. Парапофизы и диапофизы туловищных позвонков сближены. Общее количество позвонков достигает 565. Крестец изчезает. Хвостовой отдел обычно укорочен. Ребра, как правило, со слабой двухголовчатостью. Грудины нет. В клоачной области концы ребер вильчатой формы, образуя лимфапофизы (limphapophyses). Нет ни каких остатков плечевого пояса, тазовый пояс часто сохраняется. Иногда развита рудиментарная бедренная кость, снабженная когтевидным придатком (надсемейство Boidea). Голова покрыта щитками, иногда редуцированными. Тело покрыто чешуей; брюшные чешуи, как правило, расширены поперечно, образуя гастростеги; у некоторых морских змей редуцированы до полного исчезновения. То же самое можно сказать и о подхвостовых щитках. Чешуи на спине и боках тела относительно велики, часто удлинены и несут посредине продольное возвышение; иногда чешуйки расширены и имеют шестиугольную форму.
В соответствии с высоким совершенством и сложностью внутричерепной подвижности челюстная мускулатура змей достигла наивысшей степени дифференциации среди рептилий. Все основные отделы этой мускулатуры представлены сериями обособленных мышц, в целом довольно сходными у большинства змей, за исключением специализированных роющих групп (Иорданский, 1990). Огромное значение в систематике и таксономии надвидовых таксонов имеет строение и положение таких мышц, как depressor mandibulae – нижнечелюстной опускатель, adductor mandibulae externus superficialis – наружный челюстной аддуктор (верхняя порция), adductor mandibulae externus medialis – то же (средняя порция) – мышцы, приводящие в движение нижнюю челюсть (McDowell, 1967, 1972;Gopalakrishnakone, Kochva,1990).
У змей, как и у некоторых роющих ящериц, подвижные веки преобразованы в неподвижную прозрачную перепонку. Клетки сетчатки упрощены, а наружные и внутренние глазничные мышцы редуцированы. Ни у одной змеи нет маслянистых включений в светочувствительных элементах сетчатки, а у большинства – центральной ямки желтого пятна, которая имеется у продвинутых ящериц. Еще одно свидетельство дегенерации глаза – отсутствие хрящевых и окостеневших поддерживающих структур склеры, сохраняющихся даже у роющих ящериц (Walls, 1942; Kardong, 2002).
Примитивные змеи еще имеют два легких (обычно менее развито левое), в дальнейшем остается только одно. Передний конец развитого легкого функционирует как дыхательная часть, а задний гладкостенный представляет собой лишь вместилище воздуха. Некоторые змеи (в частности, морские) имеют видоизмененное строение части задней стенки трахеи: она расширяется мешкообразно, имеет губчатое строение (трахеальное легкое) и служит дополнительным местом газообмена. Желудок в виде некоторого утолщения части пищевода, плохо отграничен и лежит продольно. Печень в виде длинной лопасти. Желчный пузырь объемистый. Почки, семенники и яичники вытянуты, причем правые лежат впереди левых.
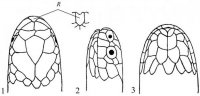
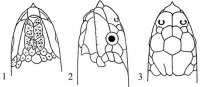
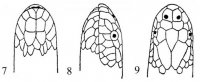
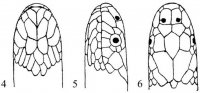
Совокупность чешуй – фолидоз (pholidosis) играет важную роль в систематике змей. Чешуи тела (squmae) могут быть гладкими, с бугорками или ребрышками, налегать друг на друга или лежать бок о бок. Крупные чешуи именуются щитками (scutae). Особенного развития последние достигли на голове (рис. I, 1-3), где покровная часть именуется пилеусом (pileus).
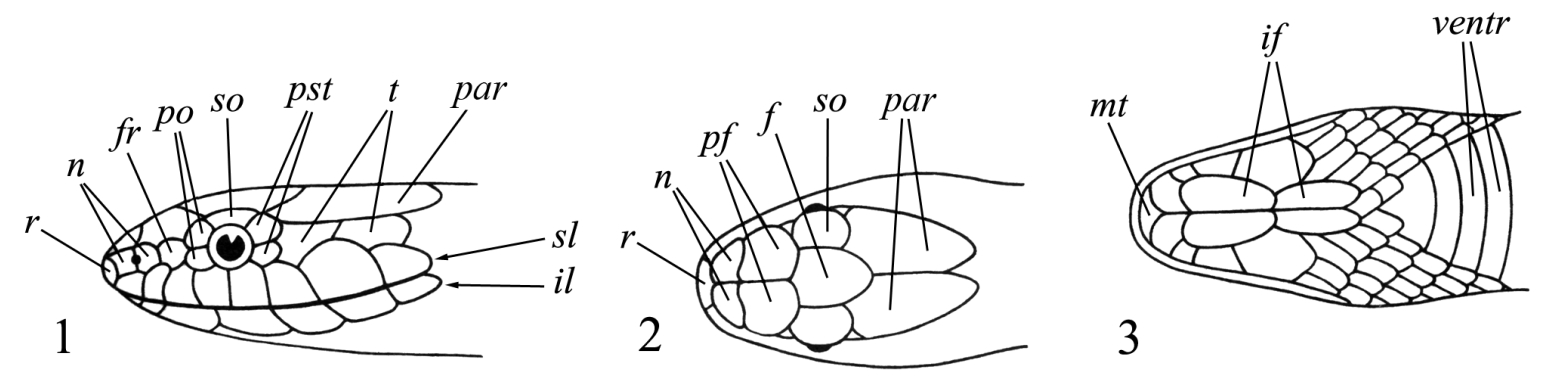
Рис. I, 1-3. Фолидоз головы змеи. Щитки: f – лобный, fr – скуловой, if – челюстные, il – нижнегубные, mt – подбородочный, n – носовой, par – теменной, pf – предлобные, po – предглазничные, pst – заглазничные, r – межчелюстной, sl – верхнегубные, so – надглазничный, t – височные, ventr – брюшные.
На ней спереди назад лежат следующие щитки: межчелюстной (praemaxillare), носовые (nasalia), предлобные (praefrontalia), лобный (frontale) и теменные (parietalia). Между носовыми могут присутствовать межносовые щитки (internasalia). На боковой стороне головы могут быть скуловой щиток (loreale= frenale), предглазничные (praeocularia), надглазничный (supraoculare), заглазничные (postocularia), височные (temporalia), верхнегубные (supralabialia) и нижнегубные (infralabialia) щитки. В месте сочленения нижних челюстей лежит подбородочный щиток (mentale), сзади которого располагаются нижнечелюстные (inframaxillaria= submaxillaria) щитки. У большинства змей нижняя поверхность тела покрыта брюшными щитками (ventralia), а хвоста – подхвостовыми щитками (subcaudalia). Между брюшными щитками и клоакой находятся один или более анальных щитков (analia). Последние могут быть сплошными или разделенными.